
Signalment: 12-month-old, male, Beagle, canine.
History: This dog arrived from a commercial vendor at approximately 5 months of age, and was clinically normal. As per our animal conditioning protocols for pretest dogs, this animal was routinely administered an anthelmintic, vaccinated, and repeated examinations of hematology, serum biochemistry, and urinalysis parameters were performed. An initial ophthalmoscopic examination was normal when the dog was 10 months old. When the dog was approximately 12 months old, both eyes appeared blue and prominent, and an ophthalmologic examination revealed bilateral buphthalmia, increased intraocular pressures, anterior uveitis, and corneal edema. A clinical diagnosis of bilateral glaucoma was made. The dog was determined to be ineligible for inclusion in safety studies and was euthanatized.
Gross Pathology: At necropsy, the eyes had blue opaque corneas. Both eyes were promptly enucleated, fixed in Zenker's fixative for several hours, rinsed in ethanol for at least two days, then sectioned at 5 mm, and stained with hematoxylin and eosin.
Laboratory Results: Hematology, serum biochemistry, and urinalysis results were within normal limits for our laboratory.
Contributor's Diagnosis and Comments: Severe bilateral granulomatous anterior uveitis.
Etiology: suspected immune-mediated response (unknown antigen).
Conference Note: Conference participants noted mild attenuation of the filtration angles, which likely caused the increased intraocular pressure by interfering with fluid drainage from the anterior chamber. However, histologic changes compatible with glaucoma (i.e. atrophy of inner retinal layers, optic disc cupping) are not present.
The pathogenesis of Vogt-Koyanagi-Harada (VKH) syndrome in humans is believed to involve the development of autoantibodies and cytotoxic T cells to melanocytes. However, human patients also develop neurologic lesions and a wide range of other immunological abnormalities. In dogs, no consistent immunological abnormalities have been observed.5
In dogs with VKH-like syndrome, uveitis usually precedes or occurs simultaneously with skin disease. The skin lesion is a symmetric facial poliosis and vitiligo, with variable erythema and crusting. Lesions may also be found on the footpads or perianal region, and in some cases can become generalized. Akitas, Samoyeds, Siberian Huskies, Alaskan Malamutes, Chow Chows, and crosses of these breeds are at increased risk.4
Another cause of uveitis and corneal edema in dogs is an immune-complex reaction to antigens of canine adenovirus 1. Uveitis in that condition is seen both in dogs with natural infection and in those vaccinated with live attenuated adenovirus vaccine. The uveitis results from the formation of virus-antibody complexes in the anterior chamber and cornea, with complement activation and consequent neutrophil accumulation. The neutrophils release enzymes that damage corneal epithelial cells, resulting in corneal edema and opacity.5 A reaction of this type is considered likely in this case.
Contributor: Merck Research Laboratories, Department of Safety Assessment, West Point, PA 19486
References:
International Veterinary Pathology Slide Bank:
Laser disc frame #4723, 4819, 10204, 16887
Signalment: 4- to 6-week old, female, BALB/c mice.
History: This group of BALB/c mice was inoculated with cowpox virus by small particle aerosol inhalation, and was sacrificed on day 12 post exposure, which in this case was the experiment's termination. Immediately prior to performing the necropsy on these mice, select tissues were collected for virus isolation and viral titer determination.
Gross Pathology: Involving 50-85% of the lungs, there
were multifocal to coalescing, sometimes lobular, areas of dark-red
consolidation. The cranial mediastinum was hemorrhagic. The thymus
was not recognizable in this group. Distal areas on the tails
had multifocal to coalescing ulcers and necrosis.
Contributor's Diagnosis and Comments: Lung: Bronchopneumonia,
necrotizing, fibrinohemorrhagic, neutrophilic and histiocytic,
multifocal to coalescing, moderate to severe, with eosinophilic
intracytoplasmic inclusion bodies, intracellular edema, and epithelial
proliferation, BALB/c mouse, rodent.
Etiology: Cowpox virus (Orthopoxvirus)
Cowpox is endemic to Europe and parts of western Asia, but despite its name, is not endemic in cattle and is seen in this species only rarely.11 Cowpox can affect a wide range of animals including wild and domestic Felidae, cattle, rodents, humans, and captive mammals to include elephants and rhinoceroses.2,11 Although the domestic cat is the most frequently reported host of cowpox virus in western Europe, the reservoir host appears to be wild rodents such as wild bank voles, field voles, and woodmice; these are species in which a high prevalence of Orthopoxvirus antibody is believed to indicate cowpox virus infection and not infection with ectromelia or vaccinia viruses.3 Domestic cats presumably become infected with cowpox virus while hunting small rodents. An increased disease attack rate during autumn in domestic cats corresponds with peak rodent population and activity. The primary cutaneous lesions seen early on in infected cats are typically ulcerative with proliferative borders and involve the head, oral cavity, neck, forelimbs and paws. Viremia, either spontaneous or related to compromised immunity, such as that seen with feline immunodeficiency virus infection, corticosteroid or progesterone therapy, results in pyrexia, and generalized cutaneous lesions. Occasionally oral and pulmonary involvement is also seen.2,11 A fibrinonecrotic, often fatal, bronchopneumonia is commonly seen in nondomestic cats, especially cheetahs.
Generally, microscopic examination also reveals involvement of the mesodermal tissues and the presence of A-type and B-type intracytoplasmic inclusion bodies; these features are present in the submitted case. A-type inclusions are large, homogeneous, eosinophilic and numerous, while B-type inclusions are somewhat faintly eosinophilic and irregularly shaped.6
Human cowpox usually results from direct cutaneous inoculation, and in endemic areas a large percentage of cases are associated with direct contact with infected rural domestic cats. Like domestic cats, human cases have a higher incidence during the late summer and early autumn in Britain and Western Europe. Cutaneous lesions in humans are usually limited to the hands and face, and systemic symptoms such as fever, malaise and lymphadenopathy are common. Death may occur in eczematous and immunocompromised individuals.1,2
Several recent findings regarding the pathogenesis of poxvirus infection are noteworthy. The poxvirus genome encodes proteins that can either antagonize or compete with molecules involved in the host-virus interaction.11 These viral proteins are members of different viral protein families consisting of the cell-associated cytokine response-modifying or serpin-related protein family, which includes a complement control protein, a neurovirulence factor and a group of cytokine-binding proteins.7
Vaccinia virus is the best characterized member of the Orthopoxvirus genus, but rodents are not natural hosts for vaccinia virus and vaccinia-infected rodents do not correlate with vaccinia or smallpox virus-infected humans. Notwithstanding, vaccinia complement control protein was the first microbial protein shown to be structurally similar to mammalian complement control proteins, to bind to C3 and C4, to block the complement cascade at a variety of sites, to block complement-dependent neutralization assays, to enhance repair of cutaneous lesions in rabbits, and to exhibit more activity than the human complement control protein counterpart. A highly conserved homolog of vaccinia complement control protein is also encoded by the small pox and cowpox viruses. Rodents are natural hosts for cowpox virus and may serve as a practical and optimal model for simulating natural smallpox virus infection in humans.7
Many poxviruses (such as vaccinia, ectromelia and cowpox viruses) have been shown to express an extracellular soluble interferon-gamma binding protein which blocks receptor-ligand binding, subsequent signal induction and anti-viral effects of interferon-gamma.10
In addition to virus-encoded host-interactive proteins (homologues of host proteins which inhibit specific host defenses), host factors also contribute to the pathogenesis of poxvirus disease. For example, not all strains of mice are equally susceptible to the lethal effects of ectromelia virus infection. Such resistance to lethal effects of mousepox is apparently partially controlled by multiple unlinked autosomal dominant genes, some of which encode the natural killer cell receptor complex and the fifth component of complement.4
Poxviruses represent a large family of double-stranded DNA viruses that infect vertebrate and invertebrate hosts. Cowpox virus belongs to the Orthopoxvirus genus of the subfamily Chordopoxvirinae, family Poxviridae. Vaccinia virus, the prototypal member of the Orthopoxvirus genus, was the first animal virus seen microscopically, cultured, accurately titered, purified, and analyzed; all orthopoxviruses are closely related and have not been demonstrated to exhibit cross-hybridization with members of other poxvirus genera.6,9 Although cowpox virus was originally used by Jenner in 1796 to debut vaccination, at that time against smallpox in humans, vaccinia virus has since been used for vaccination against smallpox.
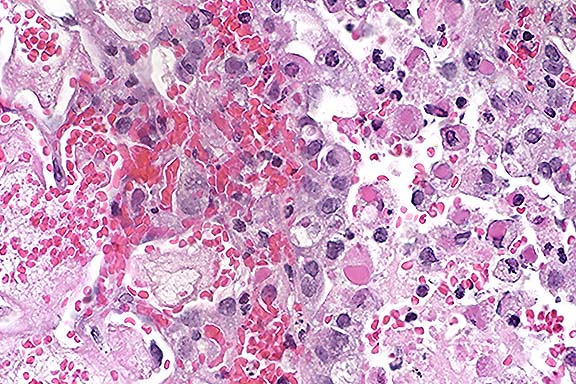
Conference Note: Tissue sections that had been immunohistochemically stained for poxviral antigen were viewed in conference. Viral antigen was demonstrated in intracytoplasmic inclusions in numerous cell types, including bronchiolar epithelial cells, alveolar macrophages and pneumocytes, and smooth muscle cells. Electron micrographs also demonstrated typical pox virions within the cytoplasm of numerous cells.
Ultrastructural features of poxvirus include (1) a nucleocapsid core (a shell of inner and outer membranes containing viral DNA in the form of linear 200,000-base pair duplex molecules with hairpin loops connecting the two strands at each end); (2) an intermediate coat (an outer membrane and an inner shell that expands into two lateral bodies); and (3) the lipoprotein envelope. A dense body is adjacent to the inner shell.12
Contributor: Pathology Division, U.S. Army Medical Research Institute of Infectious Disease, Ft. Detrick, Frederick, MD 21702-5011
References:
Signalment: 100-week-old, male, CD rat.
History: The rat was found dead in its cage near the end of a carcinogenicity study.
Gross Pathology: The right adrenal gland was enlarged (10 mm diameter). For comparison, the left adrenal gland was 5 x 4 x 4 mm.
Contributor's Diagnosis and Comments: Ganglioneuroma, adrenal, rat.

The differential diagnosis of adrenal medullary neoplasms containing a mixture of ganglion cells, neural stromal cells, and neoplastic pheochromocytes is controversial. These components most likely represent divergent differentiation from a common progenitor cell.2,3 Hamlin and Banas recommend that, due to the "phenotypic plasticity of neoplastic medullary cells", only those tumors consisting of >80% ganglion cells and nerve fibers should be diagnosed as ganglioneuromas.2
Contributor: Covance Laboratories, P.O. Box 7545, Madison, WI 53704
References:
Signalment: 12-year-old, neutered, horse.
History: This animal was necropsied in winter, and was part of a survey to study the prevalence at necropsy of taeniasis among grazing horses in France. It died from an unrelated condition.
Gross Pathology: Up to 300 parasites were attached to the mucosa of the proximal cecum, near the ileocecal junction. These parasites were identified macrosocpically as Anoplocephala perfoliata. Reddish discoloration of the mucosa with multifocal ulceration were observed.
Contributor's Diagnosis and Comments: Cecum: typhlitis, erosive, severe, locally extensive with attached Anoplocephala perfoliata, and luminal Balantidium coli.
The mucosa is ulcerated, slightly depressed, and is matted with an inflammatory exudate composed of fibrin admixed with degenerated eosinophils. A few sections of a 50 to 80 mm protozoan parasite showing a macronucleus are present superficially. They are interpreted as Balantidium coli. There is some mucoid metaplasia of the glandular epithelium. The lamina propria is infiltrated by numerous lymphocytes, plasma cells and eosinophils, with fewer macrophages. The submucosa is moderately edematous and is similarly inflitrated. It contains prominent blood vessels. A section of cestode, morphologically compatible with Anoplocephala perfoliata is observed in the lumen.
Anoplocephala perfoliata, which is among the most frequent intestinal parasites in horses, is commonly associated with lesions and occasionally with mortality. Although some horses were heavily infested in this study (sometimes several thousands of parasites), the parasite was never the cause of death.
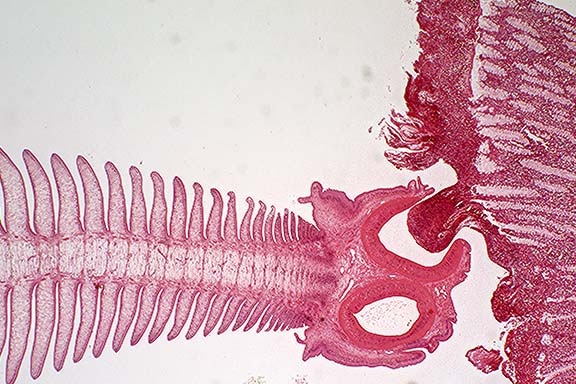
Conference Note: This case was reviewed by C. H. Gardiner, PhD, veterinary parasitology consultant to the AFIP. He believes that the genus of the ciliated protozoan cannot be definitively determined in this section. Fixed, trichrome-stained fecal samples are usually needed to identify Balantidium sp. As the contributor noted, Balantidium is usually found in primates and swine; there are numerous other genera of protozoa which may be found in the intestines of horses.
Gross lesions of Anoplocephala perfoliata infection in the horse include mucosal erosion and ulceration, especially at the ileocecal orifice. The depressed surface is often covered by a fibrinous exudate, sometimes with hemorrhage. Ileocecal and cecocecal intussusception, and occasionally intestinal perforation, have also been associated with infection by this cestode.1
Other tapeworms of horses include Anoplocephala magna and Paranoplocephala mamillana, both of which are found in the small intestine and rarely in the stomach. A. magna in large numbers can cause a catarrhal or hemorrhagic enteritis, whereas P. mamillana is seldom associated with significant lesions.3 All of these cestodes use oribatid mites as intermediate hosts. Cysticercoids are produced in these mites 2 to 4 months after infection. Horses become infected by ingesting the mites with feedstuffs, and adult worms can be found 4 to 6 weeks after ingestion of the mites.
Contributor: Pfizer, Centre de Recherches, Department of Pathology, 37401 AMBOISE, France.
References:
International Veterinary Pathology Slide Bank:
Laser disc frame #592, 1077, 2419, 14617-18, 20209, 22170
Terrell W. Blanchard
Major, VC, USA
Registry of Veterinary Pathology*
Department of Veterinary Pathology
Armed Forces Institute of Pathology
(202)782-2615; DSN: 662-2615
Internet: blanchard@email.afip.osd.mil
* The American Veterinary Medical Association and the American College of Veterinary Pathologists are co-sponsors of the Registry of Veterinary Pathology. The C.L. Davis Foundation also provides substantial support for the Registry.